Comparative Genomics Determines Strain-Dependent Secondary Metabolite Production in Streptomyces venezuelae Strains

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cell Growth
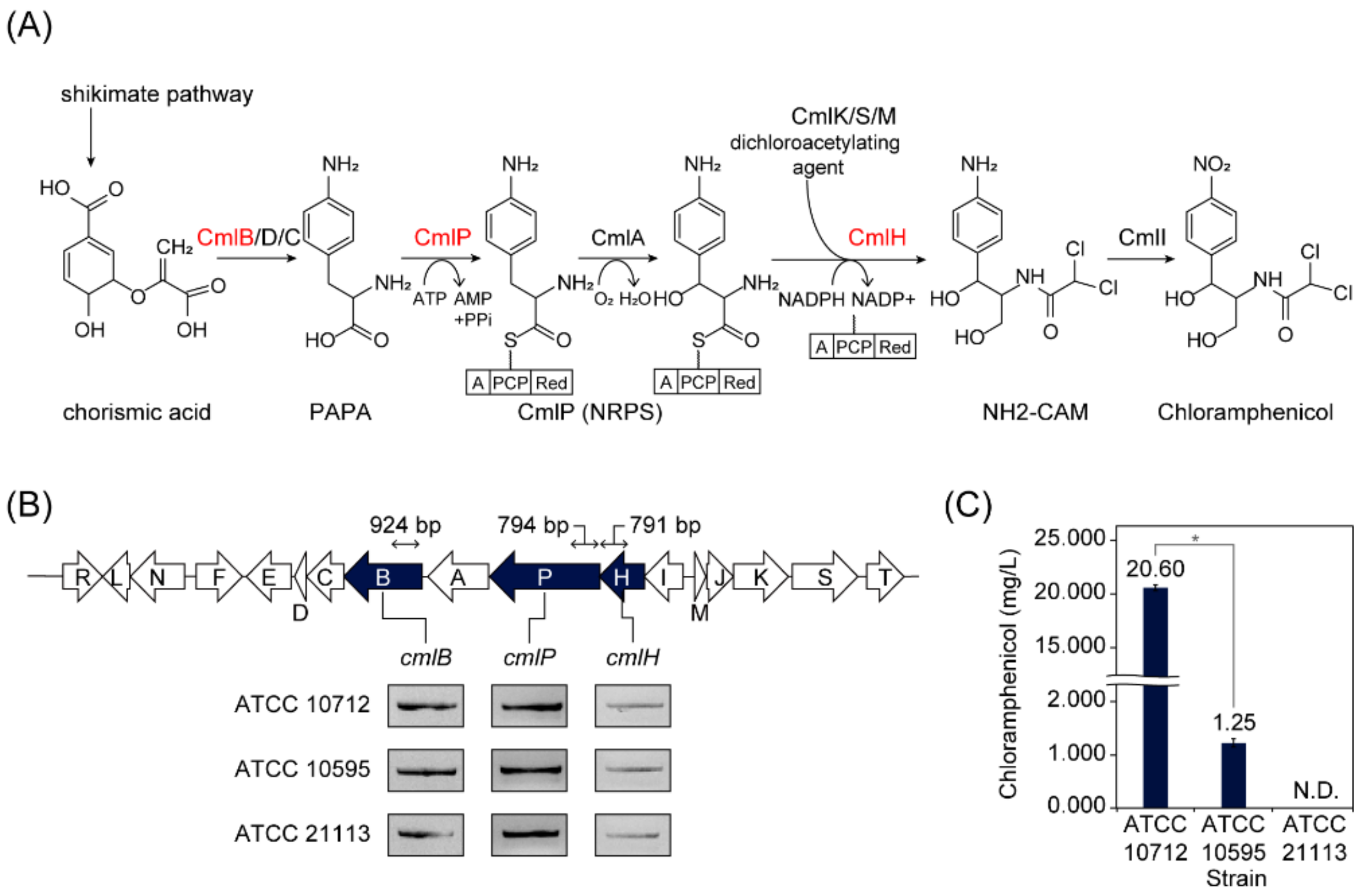
2.2. Screening S. venezuelae Strains Harboring Chloramphenicol BGCs
2.3. Genome Sequencing of Three S. venezuelae Strains
2.4. Comparative Genome Analysis
2.5. Genome Analyses
2.6. Measurement of Chloramphenicol and Jadomycin
2.7. Engineering of S. venezuelae ATCC21113 Using CRISPR/Cas9 System
3. Results
3.1. Screening S. venezuelae Strains Harboring Chloramphenicol BGC
3.2. Genome Completion of the Three S. venezuelae Strains Harboring Chloramphenicol BGC
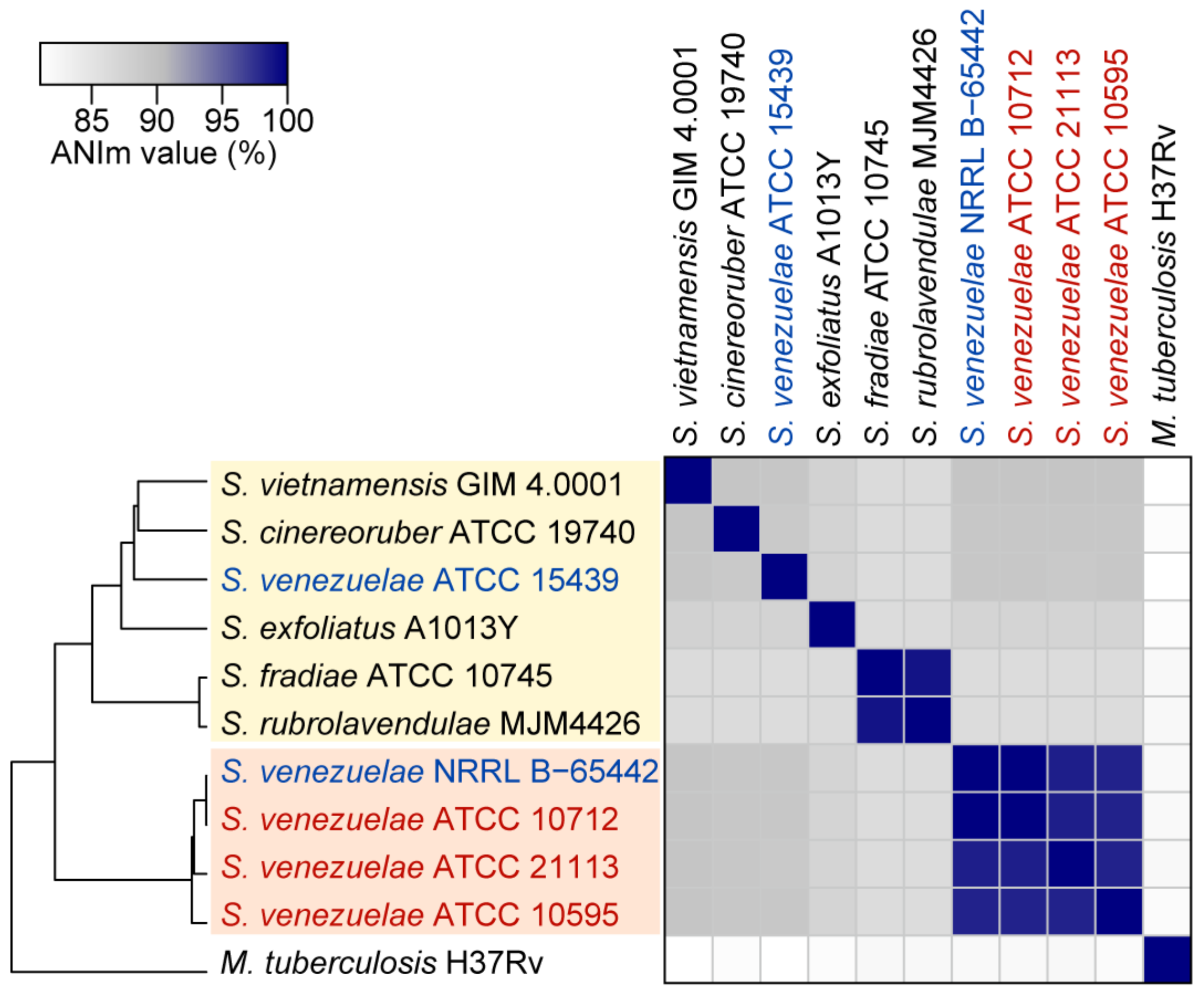
3.3. Genomic Divergence between S. venezuelae Strains
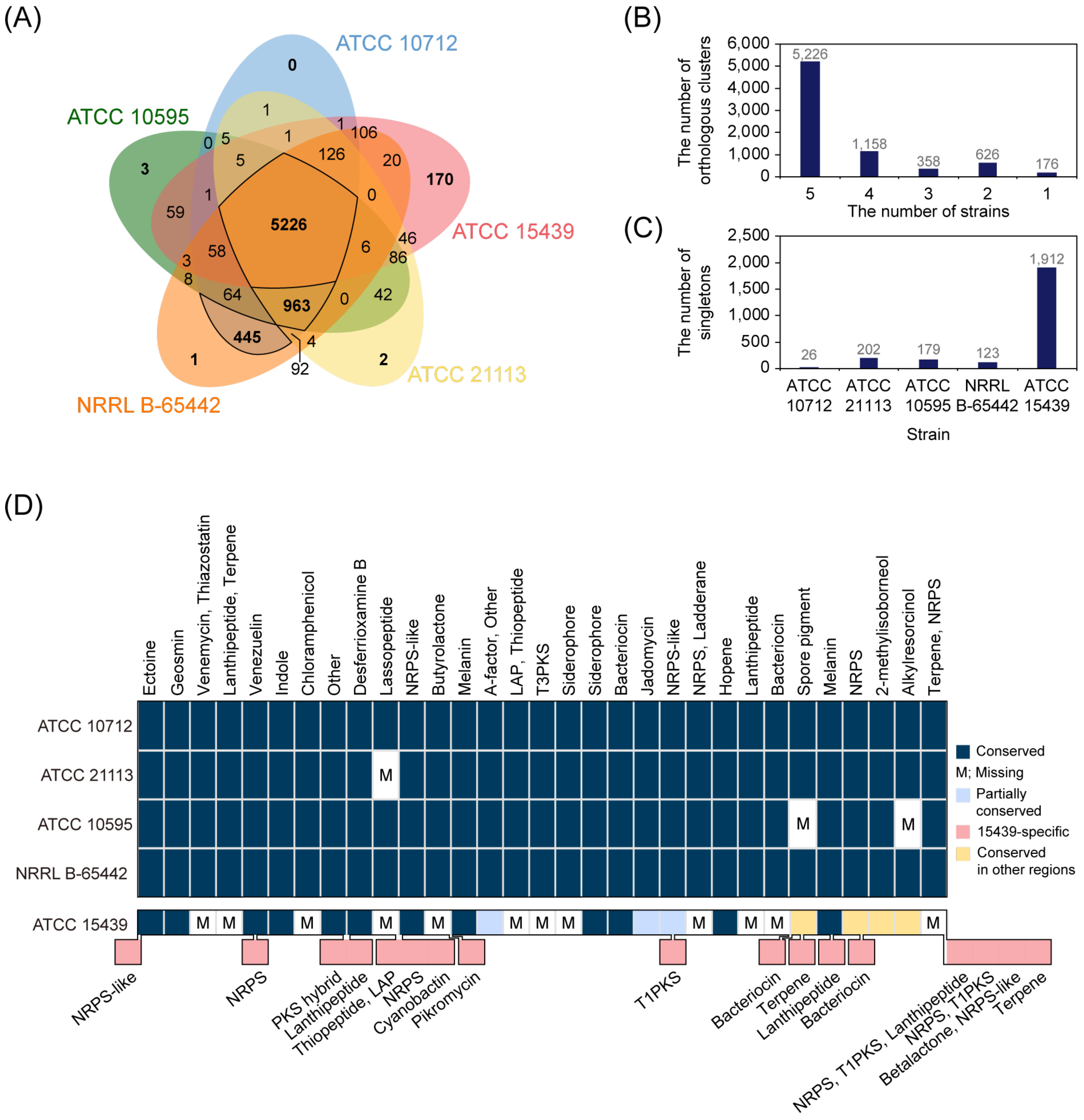
3.4. Highly Conserved smBGCs among S. venezuelae Strains
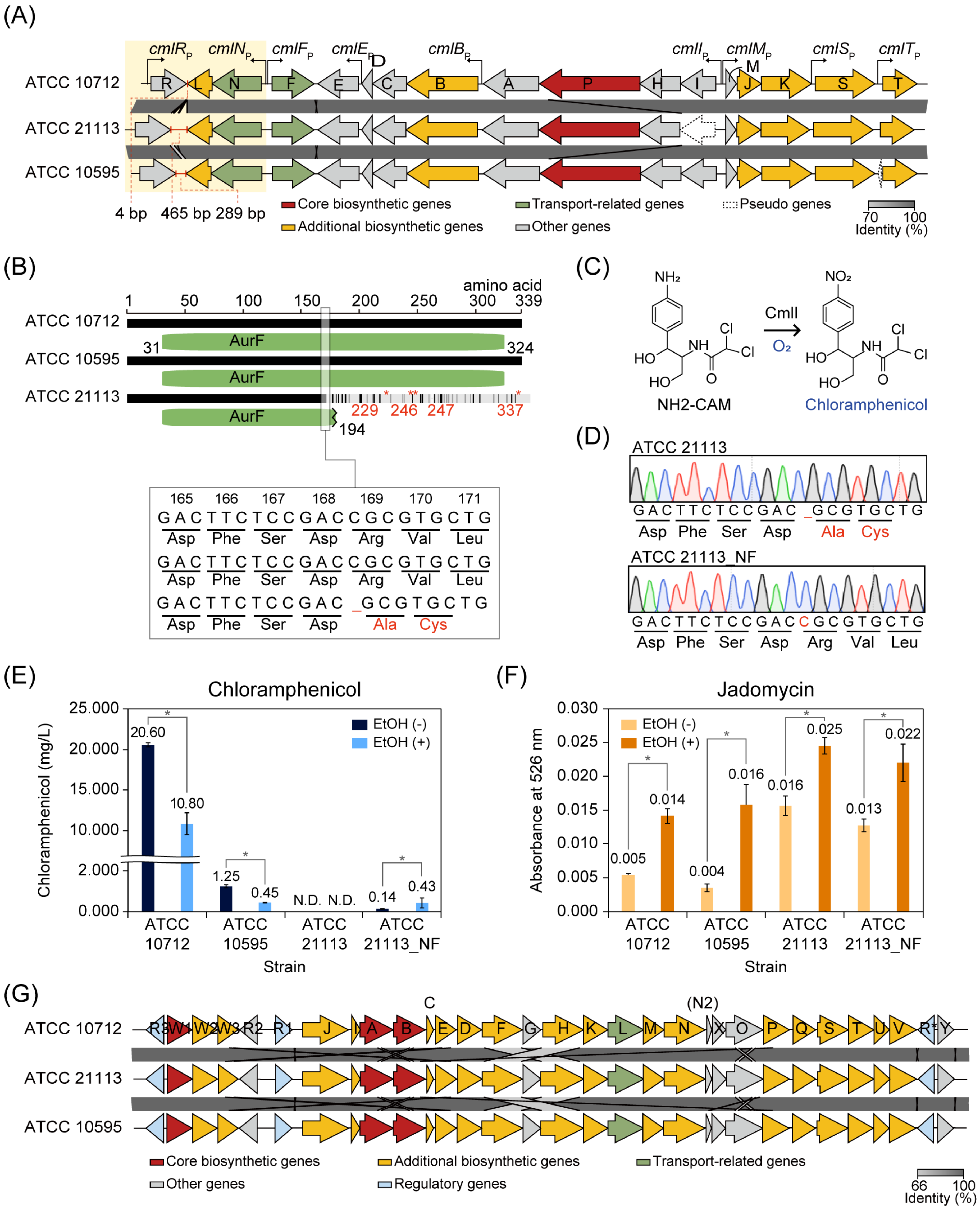
3.5. Comparison of Chloramphenicol BGCs of S. venezuelae Strains
3.6. Comparison of Jadomycin BGCs of S. venezuelae Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Demain, A.L. Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol. Biotechnol. 2014, 41, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A. Streptomyces in Nature and Medicine: The Antibiotic Makers; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Zhou, Z.; Gu, J.; Du, Y.L.; Li, Y.Q.; Wang, Y. The -omics era- toward a systems-level understanding of Streptomyces. Curr. Genom. 2011, 12, 404–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, M.J.; Tschowri, N.; Schlimpert, S.; Flardh, K.; Buttner, M.J. c-di-GMP signalling and the regulation of developmental transitions in streptomycetes. Nat. Rev. Genet. 2015, 13, 749–760. [Google Scholar] [CrossRef]
- Phelan, R.M.; Sachs, D.; Petkiewicz, S.J.; Barajas, J.F.; Blake-Hedges, J.M.; Thompson, M.G.; Reider Apel, A.; Rasor, B.J.; Katz, L.; Keasling, J.D. Development of next generation synthetic biology tools for use in Streptomyces venezuelae. ACS Synth. Biol. 2017, 6, 159–166. [Google Scholar] [CrossRef]
- Ehrlich, J.; Gottlieb, D.; Burkholder, P.R.; Anderson, L.E.; Pridham, T.G. Streptomyces venezuelae, n. sp., the source of chloromycetin. J. Bacteriol. 1948, 56, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; Wiley: Hoboken, NJ, USA, 2002. [Google Scholar]
- Doull, J.L.; Ayer, S.W.; Singh, A.K.; Thibault, P. Production of a novel polyketide antibiotic, jadomycin B, by Streptomyces venezuelae following heat shock. J. Antibiot. (Tokyo) 1993, 46, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Jakeman, D.L.; Bandi, S.; Graham, C.L.; Reid, T.R.; Wentzell, J.R.; Douglas, S.E. Antimicrobial activities of jadomycin B and structurally related analogues. Antimicrob. Agents Chemother. 2009, 53, 1245–1247. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.R.; Blundon, H.L.; Ladda, M.A.; Robertson, A.W.; Martinez-Farina, C.F.; Jakeman, D.L.; Goralski, K.B. Jadomycin breast cancer cytotoxicity is mediated by a copper-dependent, reactive oxygen species-inducing mechanism. Pharmacol. Res. Perspect. 2015, 3, e00110. [Google Scholar] [CrossRef]
- Thanapipatsiri, A.; Gomez-Escribano, J.P.; Song, L.J.; Bibb, M.J.; Al-Bassam, M.; Chandra, G.; Thamchaipenet, A.; Challis, G.L.; Bibb, M.J. Discovery of unusual biaryl polyketides by activation of a silent Streptomyces venezuelae biosynthetic gene cluster. Chembiochem 2016, 17, 2189–2198. [Google Scholar] [CrossRef] [Green Version]
- Inahashi, Y.; Zhou, S.S.; Bibb, M.J.; Song, L.J.; Al-Bassam, M.M.; Bibb, M.J.; Challis, G.L. Watasemycin biosynthesis in Streptomyces venezuelae: Thiazoline C-methylation by a type B radical-SAM methylase homologue. Chem. Sci. 2017, 8, 2823–2831. [Google Scholar] [CrossRef] [Green Version]
- Rabe, P.; Rinkel, J.; Klapschinski, T.A.; Barra, L.; Dickschat, J.S. A method for investigating the stereochemical course of terpene cyclisations. Org. Biomol. Chem. 2016, 14, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Li, B.; Claesen, J.; Shi, Y.X.; Bibb, M.J.; van der Donk, W.A. Discovery of unique lanthionine synthetases reveals new mechanistic and evolutionary insights. PLoS Biol. 2010, 8, e1000339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidda, J.D.; Song, L.J.; Poon, V.; Al-Bassam, M.; Lazos, O.; Buttner, M.J.; Challis, G.L.; Corre, C. Discovery of a family of gamma-aminobutyrate ureas via rational derepression of a silent bacterial gene cluster. Chem. Sci. 2014, 5, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Kodani, S.; Komaki, H.; Suzuki, M.; Kobayakawa, F.; Hemmi, H. Structure determination of a siderophore peucechelin from Streptomyces peucetius. Biometals 2015, 28, 791–801. [Google Scholar] [CrossRef]
- Som, N.F.; Heine, D.; Holmes, N.A.; Munnoch, J.T.; Chandra, G.; Seipke, R.F.; Hoskisson, P.A.; Wilkinson, B.; Hutchings, M.I. The conserved actinobacterial two-component system MtrAB coordinates chloramphenicol production with sporulation in Streptomyces venezuelae NRRL B-65442. Front. Microbiol. 2017, 8, 1145. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhao, L.; Liu, H.W.; Sherman, D.H. A gene cluster for macrolide antibiotic biosynthesis in Streptomyces venezuelae: Architecture of metabolic diversity. Proc. Natl. Acad. Sci. USA 1998, 95, 12111–12116. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Yang, I.; Yoon, Y.J. Developing Streptomyces venezuelae as a cell factory for the production of small molecules used in drug discovery. Arch. Pharmacal Res. 2015, 38, 1606–1616. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeno-Tarraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef]
- Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Shinose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Omura, S. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Doroghazi, J.R.; Metcalf, W.W. Comparative genomics of actinomycetes with a focus on natural product biosynthetic genes. BMC Genom. 2013, 14, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belknap, K.C.; Park, C.J.; Barth, B.M.; Andam, C.P. Genome mining of biosynthetic and chemotherapeutic gene clusters in Streptomyces bacteria. Sci. Rep. 2020, 10, 2003. [Google Scholar] [CrossRef] [PubMed]
- Varela-Alvarez, E.; Andreakis, N.; Lago-Leston, A.; Pearson, G.A.; Serrao, E.A.; Procaccini, G.; Duarte, C.M.; Marba, N. Genomic DNA isolation from green and brown algae (Caulerpales and Fucales) for microsatellite library construction. J. Phycol. 2006, 42, 741–745. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Kim, W.; Hwang, S.; Lee, Y.; Cho, S.; Palsson, B.; Cho, B.K. Thirty complete Streptomyces genome sequences for mining novel secondary metabolite biosynthetic gene clusters. Sci. Data 2020, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods-UK 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. AntiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Jakeman, D.L.; Graham, C.L.; Young, W.; Vining, L.C. Culture conditions improving the production of jadomycin B. J. Ind. Microbiol. Biotechnol. 2006, 33, 767–772. [Google Scholar] [CrossRef]
- Zheng, J.T.; Wang, S.L.; Yang, K.Q. Engineering a regulatory region of jadomycin gene cluster to improve jadomycin B production in Streptomyces venezuelae. Appl. Microbiol. Biotechnol. 2007, 76, 883–888. [Google Scholar] [CrossRef]
- Cobb, R.E.; Wang, Y.J.; Zhao, H.M. High-efficiency multiplex genome editing of Streptomyces species using an engineered CRISPR/Cas system. ACS Synth. Biol. 2015, 4, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Kieser, T.; Foundation, J.I. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Carter, H.E.; Gottlieb, D.; Anderson, H.W. Chloromycetin and streptothricin. Science 1948, 107, 113. [Google Scholar] [CrossRef]
- Brown, M.P.; Aidoo, K.A.; Vining, L.C. A role for pabAB, a p-aminobenzoate synthase gene of Streptomyces venezuelae ISP5230, in chloramphenicol biosynthesis. Microbiology 1996, 142, 1345–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacholec, M.; Sello, J.K.; Walsh, C.T.; Thomas, M.G. Formation of an aminoacyl-S-enzyme intermediate is a key step in the biosynthesis of chloramphenicol. Org. Biomol. Chem. 2007, 5, 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Magarvey, N.; Piraee, M.; Vining, L.C. The gene cluster for chloramphenicol biosynthesis in Streptomyces venezuelae ISP5230 includes novel shikimate pathway homologues and a monomodular non-ribosomal peptide synthetase gene. Microbiology 2001, 147, 2817–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Martinez, L.T.; Borsetto, C.; Gomez-Escribano, J.P.; Bibb, M.J.; Al-Bassam, M.M.; Chandra, G.; Bibb, M.J. New insights into chloramphenicol biosynthesis in Streptomyces venezuelae ATCC 10712. Antimicrob. Agents Chemother 2014, 58, 7441–7450. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Yang, K.; Kulowski, K.; Wendt-Pienkowski, E.; Hutchinson, C.R.; Vining, L.C. An acyl-coenzyme A carboxylase encoding gene associated with jadomycin biosynthesis in Streptomyces venezuelae ISP5230. Microbiology 2000, 146, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Kulowski, K.; Wendt-Pienkowski, E.; Han, L.; Yang, K.Q.; Vining, L.C.; Hutchinson, C.R. Functional characterization of the jadI gene as a cyclase forming angucyclinones. J. Am. Chem. Soc. 1999, 121, 1786–1794. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Wang, J.; Wang, L.; Tian, X.; Yang, H.; Fan, K.; Yang, K.; Tan, H. “Pseudo“ gamma-butyrolactone receptors respond to antibiotic signals to coordinate antibiotic biosynthesis. J. Biol. Chem. 2010, 285, 27440–27448. [Google Scholar] [CrossRef] [Green Version]
- Lind, A.L.; Wisecaver, J.H.; Lameiras, C.; Wiemann, P.; Palmer, J.M.; Keller, N.P.; Rodrigues, F.; Goldman, G.H.; Rokas, A. Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 2017, 15, e2003583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.A.; Rokas, A.; Slot, J.C. Horizontal transfer and death of a fungal secondary metabolic gene cluster. Genome Biol. Evol. 2012, 4, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.J.; Jasniewski, A.J.; Que, L.; Lipscomb, J.D. Diiron monooxygenases in natural product biosynthesis. Nat. Prod. Rep. 2018, 35, 646–659. [Google Scholar] [CrossRef]
- Komor, A.J.; Rivard, B.S.; Fan, R.; Guo, Y.; Que, L., Jr.; Lipscomb, J.D. CmlI N-oxygenase catalyzes the final three steps in chloramphenicol biosynthesis without dissociation of intermediates. Biochemistry 2017, 56, 4940–4950. [Google Scholar] [CrossRef] [PubMed]
- Moody, M.J.; Jones, S.E.; Crisante, D.A.; Elliot, M.A. Streptomyces bacteria: Specialized metabolism, inter-species interations and non-coding RNAs. In Non-Coding RNAs and Inter-Kingdom Communication; Springer: Berlin, Germany, 2016; pp. 83–101. [Google Scholar]
- Zhou, K.; Aertsen, A.; Michiels, C.W. The role of variable DNA tandem repeats in bacterial adaptation. FEMS Microbiol. Rev. 2014, 38, 119–141. [Google Scholar] [CrossRef] [Green Version]
- Sekurova, O.N.; Zhang, J.; Kristiansen, K.A.; Zotchev, S.B. Activation of chloramphenicol biosynthesis in Streptomyces venezuelae ATCC 10712 by ethanol shock: Insights from the promoter fusion studies. Microb. Cell Fact. 2016, 15, 85. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Niu, G.Q.; Chater, K.F.; Tian, Y.Q.; Zhang, J.H.; Tan, H.R. Specialised metabolites regulating antibiotic biosynthesis in Streptomyces spp. FEMS Microbiol. Rev. 2016, 40, 554–573. [Google Scholar] [CrossRef] [Green Version]
- Ayer, S.W.; Mcinnes, A.G.; Thibault, P.; Walter, J.A.; Doull, J.L.; Parnell, T.; Vining, L.C. Jadomycin, a novel 8H-benz[b]oxazolo[3,2-f]phenanthridine antibiotic from Streptomyces venezuelae ISP5230. Tetrahedron Lett. 1991, 32, 6301–6304. [Google Scholar] [CrossRef]
- Doull, J.L.; Singh, A.K.; Hoare, M.; Ayer, S.W. Conditions for the production of jadomycin B by Streptomyces venezuelae ISP5230: Ecffects of heat shock, ethanol treatment and phage infection. J. Ind. Microbiol. 1994, 13, 120–125. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zou, Z.Z.; Niu, G.Q.; Tan, H.R. JadR* and jadR2 act synergistically to repress jadomycin biosynthesis. Sci. China Life Sci. 2013, 56, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Du, D.; Zhang, Y.; Zhang, J.; Niu, G.; Tan, H. A gamma-butyrolactone-sensing activator/repressor, JadR3, controls a regulatory mini-network for jadomycin biosynthesis. Mol. Microbiol. 2014, 94, 490–505. [Google Scholar] [CrossRef] [PubMed]
- Doroghazi, J.R.; Buckley, D.H. Intraspecies comparison of Streptomyces pratensis genomes reveals high levels of recombination and gene conservation between strains of disparate geographic origin. BMC Genom. 2014, 15, 970. [Google Scholar] [CrossRef] [Green Version]
- Komaki, H.; Tamura, T. Reclassification of Streptomyces rimosus subsp. paromomycinus as Streptomyces paromomycinus sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 2577–2583. [Google Scholar] [CrossRef] [PubMed]
- Saygin, H.; Ay, H.; Guven, K.; Cetin, D.; Sahin, N. Streptomyces cahuitamycinicus sp. nov., isolated from desert soil and reclassification of Streptomyces galilaeus as a later heterotypic synonym of Streptomyces bobili. Int. J. Syst. Evol. Microbiol. 2020, 70, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Sottorff, I.; Wiese, J.; Lipfert, M.; Preusske, N.; Sonnichsen, F.D.; Imhoff, J.E. Different secondary metabolite profiles of phylogenetically almost identical Streptomyces griseus strains originating from geographically remote locations. Microorganisms 2019, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- DePamphilis, M.L. DNA Replication in Eukaryotic Cells; Cold Spring Harbor Laboratory: Suffolk, NY, USA, 1996. [Google Scholar]
- Cho, H.S.; Jo, J.C.; Shin, C.H.; Lee, N.; Choi, J.S.; Cho, B.K.; Roe, J.H.; Kim, C.W.; Kwon, H.J.; Yoon, Y.J. Improved production of clavulanic acid by reverse engineering and overexpression of the regulatory genes in an industrial Streptomyces clavuligerus strain. J. Ind. Microbiol. Biotechnol. 2019, 46, 1205–1215. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | ATCC 10712 | ATCC 10595 | ATCC 21113 |
|---|---|---|---|
| Complete genome size (bp) | 8,223,505 | 7,871,480 | 7,893,803 |
| G + C content (%) | 72.5 | 72.5 | 72.5 |
| CDS | 7377 | 6942 | 6987 |
| rRNAs | 21 | 21 | 21 |
| tRNAs | 67 | 67 | 67 |
| Origin | Venezuela: Caracas, soil | USA: Illinois, compost soil | Unknown |
| Predicted secondary metabolite biosynthetic gene clusters | 30 | 27 | 30 |
| Accession No. | CP029197 | CP029195 | CP029196 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Lee, N.; Hwang, S.; Lee, Y.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.-K. Comparative Genomics Determines Strain-Dependent Secondary Metabolite Production in Streptomyces venezuelae Strains. Biomolecules 2020, 10, 864. https://doi.org/10.3390/biom10060864
Kim W, Lee N, Hwang S, Lee Y, Kim J, Cho S, Palsson B, Cho B-K. Comparative Genomics Determines Strain-Dependent Secondary Metabolite Production in Streptomyces venezuelae Strains. Biomolecules. 2020; 10(6):864. https://doi.org/10.3390/biom10060864
Chicago/Turabian StyleKim, Woori, Namil Lee, Soonkyu Hwang, Yongjae Lee, Jihun Kim, Suhyung Cho, Bernhard Palsson, and Byung-Kwan Cho. 2020. "Comparative Genomics Determines Strain-Dependent Secondary Metabolite Production in Streptomyces venezuelae Strains" Biomolecules 10, no. 6: 864. https://doi.org/10.3390/biom10060864